E’ sera, apro il portatile e inizio a sfogliare vecchie fotografie…
Non sono così vecchie. In fondo sono solamente di 15 – 13 anni fa. Esplorazioni in Valsolda e Val Sanagra, grotta della Noga, ricerche di fossili rimaneggiati dalle piene del fiume Po, Corna Piana, riunioni tra bloggers… Siamo sorridenti, insieme ad alcuni amici…
Il Tessi con la sua Renault dal parabrezza costantemente appannato! Andavamo a caccia di fossili sui ghiaioni e sulle prime cenge della Corna Piana.
Ogni volta che trovo una galleria di immagini di un giorno passato con lui mi si stampa un sorriso in faccia che irrompe nella tristezza di un addio che non ho detto e non posso più dirgli.
Mentre cerco foto nei meandri del mio computer, mi imbatto anche nelle immagini di famiglia. Vedo quelle delle gite con mio padre ultraottantenne che oramai posso vedere solamente in fotografia. trovo il mio ultimo nipotino piccino e grazioso mentre costruisce torri coi mattoncini e invece pochi giorni fa gli ho portato la macchina che era di mio padre. Gli serve per andare al lavoro. Non è più piccino. Ha preso la patente pochi giorni fa e mi ha portato a fare un giro mentre ero tempestato dalle emozioni. E’ in età da università, anche se lui preferisce lavorare.
L’università, le scienze, i “sassi”, il mio vecchio lavoro di preparatore di fossili. E’ per questi motivi che ho incontrato il Tessi.
Mi piace pensare di averlo scovato come una pagliuzza d’oro in un fiume, senza avere attrezzi specifici. Un grandissimo colpo di fortuna.
Da parecchi anni non frequento più l’università, ma ci ho passato a tempi alterni circa ventisette anni della mia vita tra studio e lavoro. In tutto quel tempo ho sempre amato stare con gli studenti.
Mi piaceva chiacchierare e passare del tempo con quelli che studiavano, che non spettegolavano degli altri ma che parlavano di se stessi e di quel che facevano. Stavo con gli studenti che mostravano una certa serietà, tanta passione e dedizione per le loro attività. Mi son sentito sempre un po’ studente anche io, fino a che ho chiuso per l’ultima volta alle mie spalle la porta del dipartimento di Scienze della Terra.
Andrea era anche così, più una serie di sfaccettature che lo rendevano unico. Me lo ricordo estremamente meticoloso nelle sue attività, buono e delicato, ma era anche un provocatore! Con la sua finezza e arguzia, già dopo quattro chiacchiere con lui, sentivi l’esigenza di cercare di migliorarti e ampliare i tuoi punti di vista.
E’ stato automatico trovarsi a gironzolare, esplorare, divulgare, dividere pause in dipartimento e aprirgli le porte di questo blog con totale fiducia.
Ora arrivo al punto. Erano dieci o undici anni che non ci vedevamo io e il Tessi, così come con altre persone che porto nel cuore.
Anche questo sito, questo vecchio enorme lavoro porto nel cuore, ma con questo è semplice: pago e sopravvive, non pago ed è finito. E’ solamente un sito. Quando finirà respirerò comunque, magari anche meglio di prima.
Perdendo il Tessi invece, il cuore si tormenterà sempre almeno un po’ per non avere avuto un tempo infinito da condividere e per non averlo salutato ogni volta come se fosse stata l’ultima volta. Per questo chiedo scusa ad Andrea per essere stato ignaro e completamente lontano dal dividere anche solo un briciolo di dolore della sua malattia e scusa anche al mio cuore che avrà un altro angolo ridotto a pezzetti.
Le scuse però ormai non servono a nulla. Non si può tornare in dietro, ma posso essere grato di avere avuto la fortuna di incontrarlo, di conoscerlo un po’, di percorrere un po’ di strada con lui.
Anche voi cari lettori e lettrici avete avuto la fortuna di incontrarlo qui (o potete farlo adesso cliccando!) e beh! Io sono stato più fortunato.
Come mi scriveva una manciata di ore fa l’amico Sam:”Non possiamo dimenticare i bei momenti passati insieme durante le nostre avventure”…
L’affetto dimostrato da una moltitudine di persone in questi giorni è anche il mio e di certo quello dei miei vecchi amici collaboratori di questo sito.
Il Tessi me lo ricordo ragazzo. Ora aveva quarant’anni e io cinquanta e non posso immaginare quello che ha passato. Sono impotente. Non ho parole all’altezza.
Abbraccio forte tutti i suoi famigliari e tutte le persone care che gli sono state vicino.
Ecco la mia analisi, totalmente personale e limitata alle mie conoscenze, che riguarda l’utilizzo della legna da ardere nel riscaldamento domestico.
Una bella discarica da ripulire prima di cominciare a tagliare legna
Non sarà scienza esatta perché non supportata da dati sperimentali misurati, verificati, oggettivi e ripetibili, ma mi atterrò semplicemente all’analisi di un fenomeno da alcuni punti di vista e alla formulazione di una teoria.
L’argomento mi interessa da vicino perché uno dei miei hobby è proprio quello di raccogliere, tagliare e spaccare legna ed è anche una pratica che mi fa risparmiare soldi nel riscaldamento domestico. Mi dà il buon umore la vista della fiamma nel mio moderno ed efficiente camino e amo quella vita un po’ selvaggia che deriva dallo stare all’aperto a far legna.
Così il mondo è più bello!!!
Le mie priorità e i miei valori non mi permetterebbero di bruciare legna per riscaldarmi se non lo ritenessi a basso impatto ambientale. So bene che tutto quello che faccio ha un impatto (così come tutto quello che fate voi lettori carissimi) e so che non sono un santo, ma le mie decisioni, abitudini e cambiamenti sono volti ad un impatto ambientale sempre più basso e ad un continuo miglioramento ambientale e possibilmente personale.
Dopo aver sottolineato il piacere che mi dà la vita selvaggia, la vista della fiamma e il risparmio economico, arriviamo alle prime e più ovvie problematiche e classiche osservazioni che riguardano le polveri prodotte da stufe e camini e il danno alla qualità dell’aria circostante.
E’ un dato di fatto che bruciare legna immetta scorie solide e sottili nell’aria e la combustione di metano o gpl da riscaldamento produca quasi solamente vapore acqueo, anidride carbonica e pochi gas davvero nocivi.
Questa però è la visione di chi vuole l’aria pulita a “casa sua” e quel che succede altrove non lo riguarda o non la riguarda!
Una quercia da potare e sistemare in po’!
Come sanno i miei vecchi lettori, sono molto interessato alle dinamiche del pianeta Terra e in queste dinamiche rientrano anche quelle che riguardano il ritrovamento, l’estrazione, la raffinazione, la distribuzione, la vendita, l’utilizzo finale dei combustibili fossili e la gestione della questione da parte della società.
Se siete persone intuitive, avete già capito che l’impatto dell’utilizzo dei gas dalla combustione “pulita” non si limita all’effetto della fiamma della caldaia.
Credo che ci vorrebbe un libro per descrivere gli aspetti che riguardano l’utilizzo dei combustibili fossili ai quali ho pensato in questi ultimi anni, ma vi elencherò solamente qualcosa sperando di intrigarvi. Potrei partire considerando l’impatto della costruzione di una caldaia, ma partiamo dalla “fonte” di approvigionamento…
E poi c’è da lavorare!
Per trovare giacimenti è stata “percorsa” tanta strada dal punto di vista scientifico. E’ stata sfruttata molto la ricerca di base che ci sarebbe stata comunque. Scoperti però i primi sistemi con cui svelare le posizioni dei giacimenti, la ricerca degli idrocarburi è diventata protagonista nel campo delle scienze geologiche.
Da tempo penso che tutte le branche della geologia sopravvivano solamente perché esiste la ricerca degli idrocarburi!
Non si offendano vulcanologi, glaciologi, paleontologi, ecc, ecc, ma senza Tirannosauri la “benzina” si vende ugualmente e se i Campi Flegrei esploderanno o qualche altro paese finirà sotto frane e alluvioni, la “benzina” continuerà a farla da padrone finché ci sarà!
Anche la ricerca scientifica, in qualche modo costa al nostro pianeta. Avrete capito che non sto parlando di soldi. Parlo di costi ambientali, ma questo è niente!!!
Le tempeste estive hanno fornito materiale in abbondanza…
Una volta trovati i giacimenti di combustibili fossili, andiamo ad estrarre!
Quando parlo con le persone dell’enorme impatto che ha l’estrazione del petrolio, del metano o del carbone, ecc, nella maggior parte dei casi sono certo che pensino all’inquinamento dovuto ai pozzi di estrazione, ad eventuali fughe di petrolio in mare o sulla terra ferma, incendi, residui abbandonati delle vecchie torri di estrazione, che sono certamente importanti e da considerare.
In realtà però mi riferisco alla nascita e al grandissimo sviluppo di città che prima non esistevano e che ruotano intorno al lavoro di estrazione e ai conseguenti guadagni. Case, palazzi, industrie , turismo, ma anche asili, scuole fino all’università alle quali vanno forniti insegnanti, negozi, grandi magazzini, aeroporti, ricchezza fuori scala e conseguenti follie, magari in pieno deserto, dove innaffiare una pianta sul davanzale o addirittura avere un giardino è diventato normale e possibile solamente con grandissimo dispendio di energia e ci sarebbe molto altro da aggiungere…
La raffinazione e la distribuzione dei combustibili fossili le metto insieme, anche se Petrolio e gas metano vengono gestiti e distribuiti diversamente. La raffinazione, se gestita con cura (utopia?), è la parte del processo che mi sembra meno dannosa per l’ambiente, a parte per le quantità gigantesche di materia prima che vengono lavorate, processate e il conseguente abuso di spazi, risorse ed energia.
La distribuzione invece la classifico come estremamente impattante. Noi esseri umani siamo distribuiti in ogni angolo della Terra ed è ormai ritenuto un diritto avere molte comodità a portata di mano, quindi c’è una rete di distribuzione estremamente efficiente ma altrettanto impattante per l’ambiente in cui si trova e nel quale si è scavato ovunque. Abbiamo distribuito centinaia di migliaia di chilometri di tubi più o meno piccoli o giganteschi, navigano navi enormi e discutibili nei mari di tutto il pianeta e poi sì, il resto viaggia anche in strada o su rotaia…
Arriviamo alla vendita del metano e in generale dei gas o gasoli da riscaldamento… Qui gira l’economia mondiale. L’energia per la società è tutto.
Con l’energia si fa tutto: si coltiva, si produce, si cresce industrialmente, si studiano nuove tecnologie, si cresce economicamente, aumenta la popolazione che produce più ricchezza per chi riesce a prendersela.
La gestione finale poi è un intrico di normative, burocrazia a non finire, regole e regolette e un’industria del riscaldamento che è certamente all’avanguardia, ma che ha come fine primario il guadagno economico di vari settori e forse, solo come fine secondario, la sicurezza degli impianti e un minore impatto ambientale che mi pare quanto mai falso.
A meno che non siano stupidi, i gestori dell’energia non hanno come fine primario il basso impatto ambientale e qui arrivo all’ultimo punto che mi fa stare ancora meglio con me stesso quando controllo una zona infestata da alberi esotici e invasivi, o quando taglio o poto i miei salici, o le querce di un mio vicino…
Parlo di manipolazione della verità da parte di chi gestisce le fonti di energia.
Parlo di certi prodotti industriali che vengono venduti alla massa definendoli a “impatto zero” e parlo poi delle guerre per accaparrarsi le risorse primarie o le aree attraverso le quali avviene il trasporto dei combustibili attraverso rotte navali o gasdotti.
La prima cosa che ho fatto quando è stato ufficializzato l’attacco all’Ucraina da parte della Russia è stato sì arrabbiarmi e indignarmi che nel 2023 si potesse ancora cominciare una guerra “tradizionale”, (e purtroppo anche nel 2024) ma pochi minuti dopo sono andato a spegnere la caldaia per non dare più soldi a chi vendeva il metano e comunque, quasi tutti ce la siamo fatta sotto perché non sapevamo se avremmo potuto accendere la caldaia fino alla fine dell’inverno e in quelli successivi.
Con i costi ambientali di una guerra come quella in cui ,tra le altre cose, il metano è usato come ricatto, la mia motosega, lo spacca tronchi, la mia nuova sega circolare e le polveri della mia stufa e del mio camino le trovo ancor più pulite di prima.
C’è bisogno che scriva anche che mentre un legno brucia nella mia stufa, migliaia di migliaia di tonnellate di legna crescono nei boschi?
Non ce n’è per tutti, ovvio. Siamo cresciuti troppo grazie allo sfruttamento dei combustibili fossili e ora ne paghiamo le conseguenze. Abbiamo appena iniziato a farlo.
Questo per dire che qualora la legge mi impedirà di accendere la mia stufa, mi sentirò privato di un’altra fetta di libertà.
Ribadisco il concetto di libertà. Con il riscaldamento a legna non sporco l’aria più che con quello a metano, anzi, potrebbe essere proprio il contrario.
Tutto sta a pensare se voglio la mia casa pulita e il mondo attorno sporco, o un mondo un po’ meno sporco per tutti.
La fatica poi, in un modo o nell’altro, non manca mai e tutto si può migliorare!
Sono indeciso se aprire i commenti a questo articolo. Mi piacerebbe perché non sono convinto di avere la verità in tasca e tanti punti di vista diversi aiutano a migliorare. Quel che ho scritto però mi sembra ragionevole.
Sto pensando anche allo sfruttamento dell’energia solare dal punto di vista elettrico, fotovoltaico e non solamente fotosintetico, ma anche questo ha un grosso impatto.
A livello divulgativo rispolvero qualche concetto geologico che spero possa essere utile ai frequentatori di vari ambienti outdoor.
Sarà più che altro una raccolta di immagini, appunti, links che facciano da filo conduttore all’ex divulgatore scientifico non aggiornato e fuori allenamento che è in me!
Ovviamente è solo una traccia e tanti concetti che qui mancano li spiegherò a voce durante la lezione che mi è stata richiesta dagli amici del CAI Edelweiss di Milano e sfrutterò le immagini, i gesti, le domande che mi verranno poste… Una parte importante poi verrà trattata durante l’uscita in ambiente.
Le parti scritte a parole servono giusto per chi voglia ripassare dopo aver seguito la lezione o per chi voglia curiosare un po’, ma giusto quello. Durante la presentazione utilizzerò solamente immagini e video. Molte immagini sono tratte dal libro di Alfonso Bosellini “Storia geologica d’Italia”.
Il tempo
Cominciamo dalla CLESSIDRA naturale in fotografia. Indica il primo concetto, ossia il passare del tempo che governa lento e inesorabile i processi geologici e tutti i cambiamenti in atto ogni istante sul pianeta Terra.
Lo studio della Terra non può esulare dalla conoscenza del tempo. La terra ha iniziato la sua storia con la sua massa più o meno attuale circa 4540Ma (4,54 miliardi di anni fa).
I movimenti della Terra nello spazio
La terra cambia anche la sua posizione nello spazio e lo fa molto velocemente:
V equatoriale: 1667 Km/h
V a 45° di latitudine: 1179 Km/h
V intorno al sole media: 107240,4 Km/h
L’orbita terrestre è leggermente eccentrica, l’asse terrestre ogni 25800 anni circa compie una rotazione completa (precessione degli equinozi) senza variare di troppo la sua inclinazione ma solo con piccole variazioni cicliche (nutazione) e ne consegue un’inversione delle stagioni ogni 12000 anni circa…
…ma anche il sole si muove e cambia posizione rispetto alle altre stelle, galassie, ecc e noi lo seguiamo. Non so a che velocità, ma deve essere mooolto alta e se volete fate pure le vostre ricerche!
Da alcuni movimenti intorno al sole dipendono certe tendenze climatiche terrestri che influiscono sui fenomeni erosivi, e ne faremo ancora cenno più tardi…
La Terra quindi cambia continuamente posizione nello spazio, temperatura, umidità e cambia sulla sua superficie dove noi viviamo.
Ma chi è il responsabile di questi cambiamenti superficiali?
Il “motore” della Terra
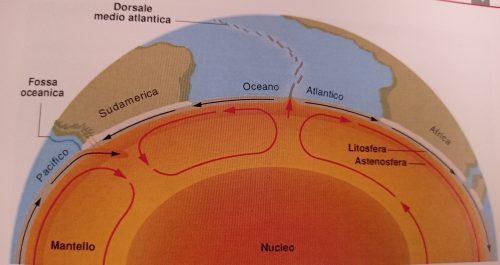
Il responsabile è l’energia interna della Terra la quale deriva in buona parte dalle reazioni di decadimento radioattivo di elementi come Uranio, Torio e Potassio, contenuti nella crosta terrestre e nel mantello.
Vediamo nelle prossime immagini tratte da “Storia geologica d’Italia” di Alfonso Bosellini:
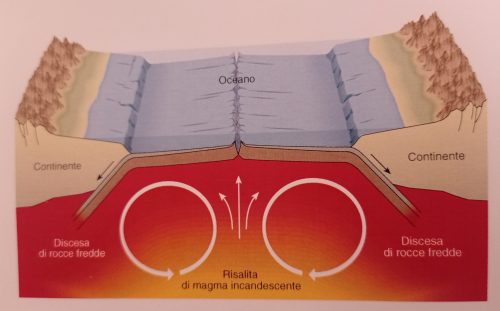
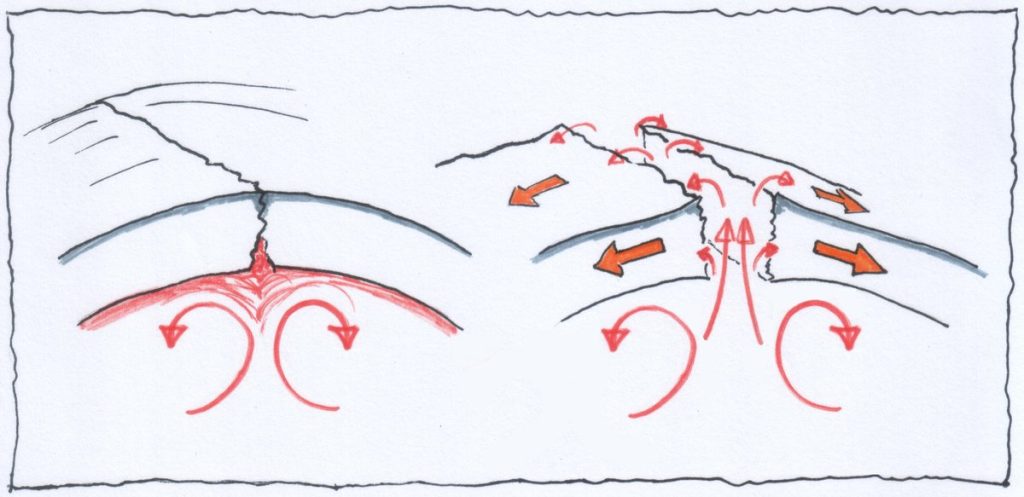
Nel mantello il calore prodotto si distribuisce per convezione, così come succede per l’acqua in ebollizione contenuta in una pentola!
Sono questi moti convettivi i responsabili della maggior parte dei movimenti orizzontali della crosta terrestre.
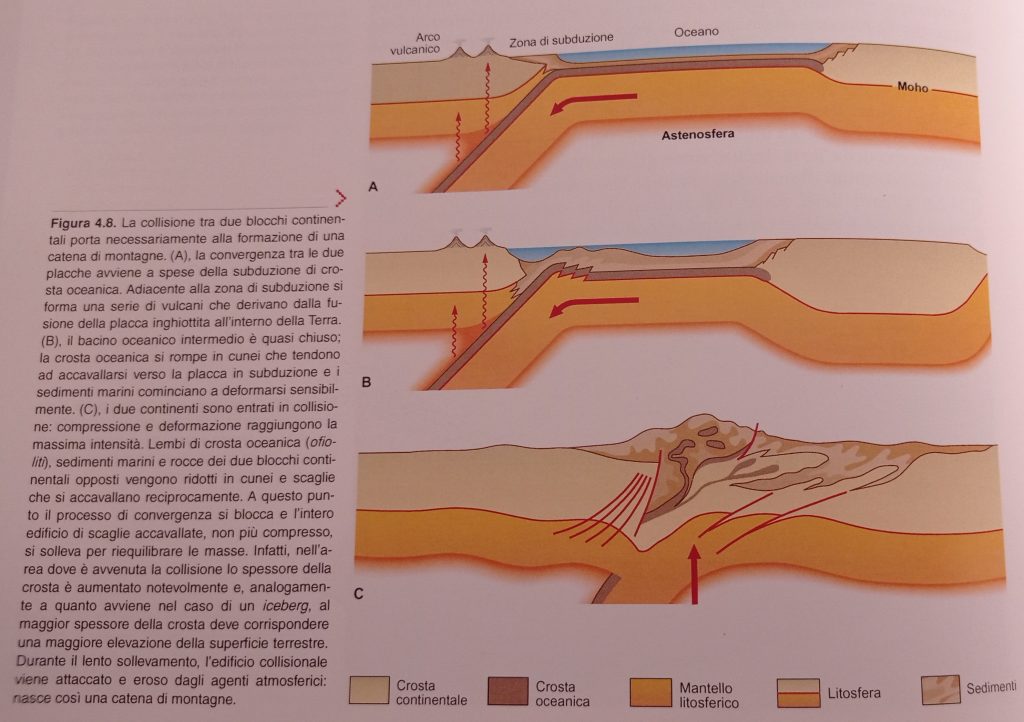
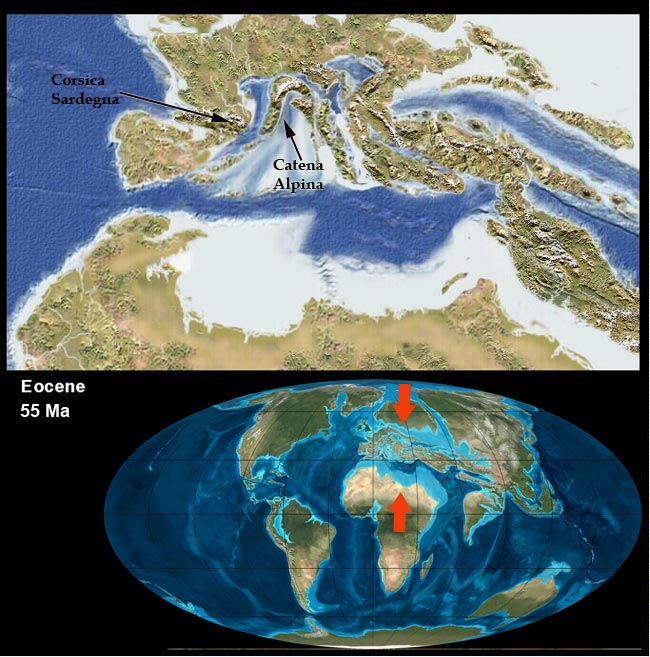
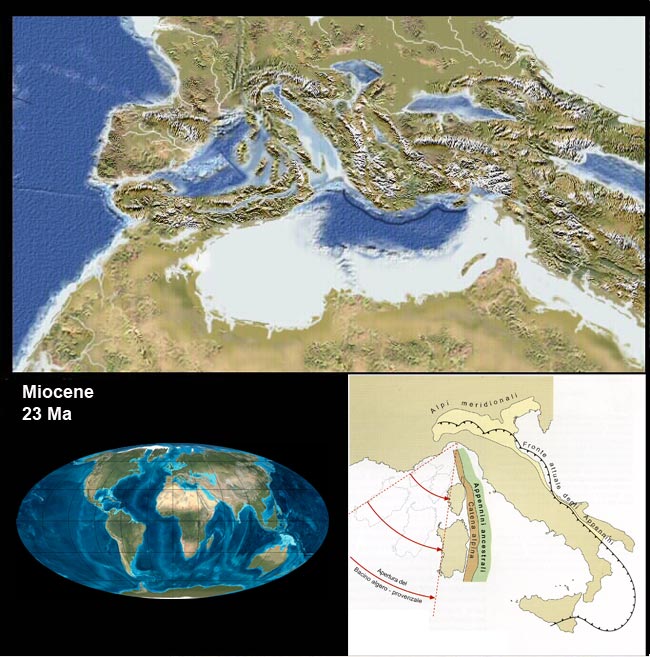
Vediamo poi in questo bel filmato (che qualche anno fa era pionieristico!) una simulazione di uno scontro tra una placca oceanica ed una continentale con conseguente subduzione della crosta oceanica, poi la successiva collisione tra placche continentali con la formazione di un rilievo come potrebbe essere stato per la catena alpina (al minuto 1′ : 25″ inizia la convergenza):
Il tutto è riassunto in tre passaggi nell’immagine successiva (sempre da Bosellini):
Come specificato manca l’erosione ed ogni processo gravitativo a modellare di continuo il tutto, ma aggiungo anche la dissoluzione delle rocce carbonatiche e non meno importante l’isostasia e le eruzioni di vulcani orogenici e anorogenici, gli impatti meteorici, ecc.
In questo video successivo una rappresentazione schematica della formazione di una rete idrica a modellare il territorio:
Molte altre tipologie di movimenti crostali sono state modellizzate. Ad esempio:
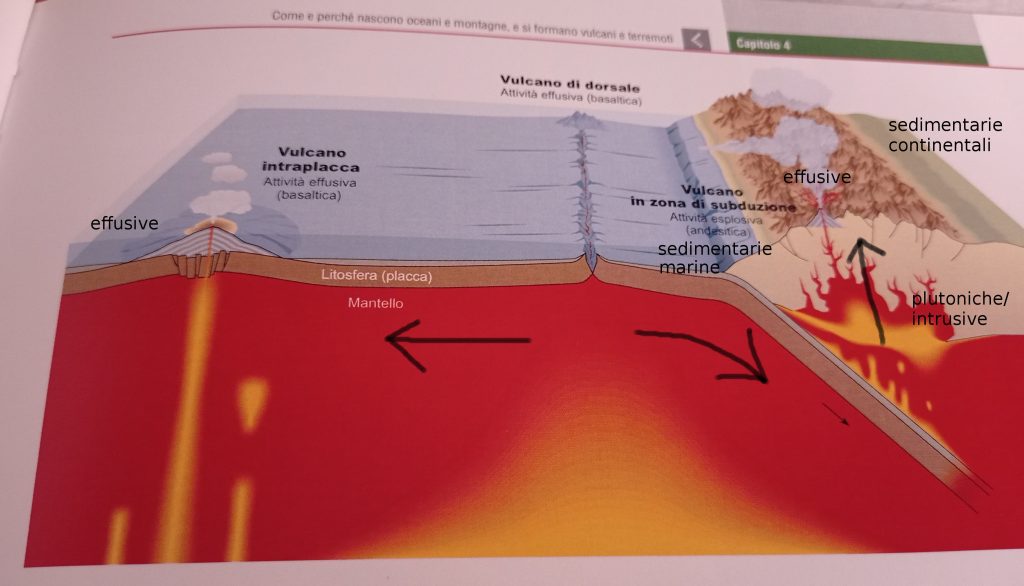
In questa immagine vediamo le principali zone di formazione delle rocce: Per le rocce ignee o magmatiche abbiamo quelle effusive del vulcanismo intraplacca, quelle effusive della zona di subduzione e quelle intrusive (o plutoniche).
Vediamo le zone di formazione delle rocce sedimentarie di origine marina e sedimentarie di origine continentale.
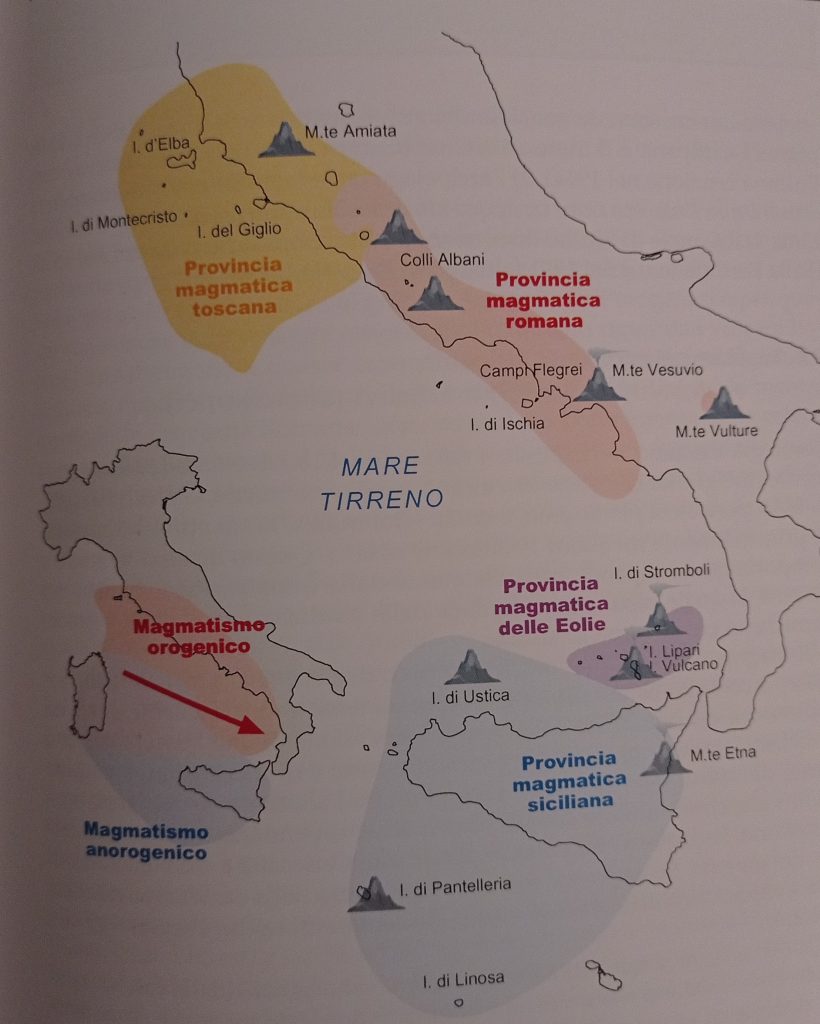
E le province magmatiche italiane dove si formano attualmente rocce ignee:
Le rocce metamorfiche caratterizzate da una trasformazione dovuta a pressione e temperatura si formano dove sia presente alta pressione e temperatura nelle zone di convergenza o nelle zone limitrofe alle intrusioni magmatiche che cedono calore alle preesistenti rocce incassanti le quali tenderanno a trasformarsi.
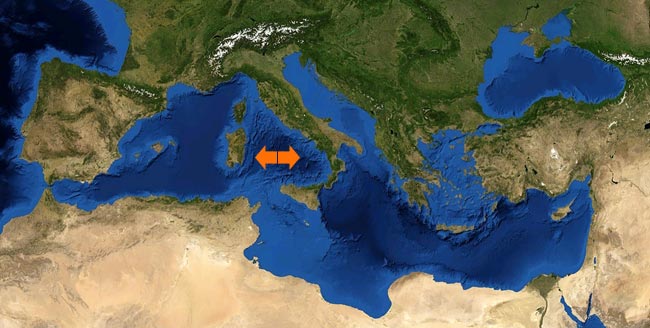
uno sguardo all’Italia e alla sua storia
Mari, montagne, pianure, fiumi, laghi e chi più ne ha più ne metta…
Galleria lez. geo. Edelweiss-003-tettonica
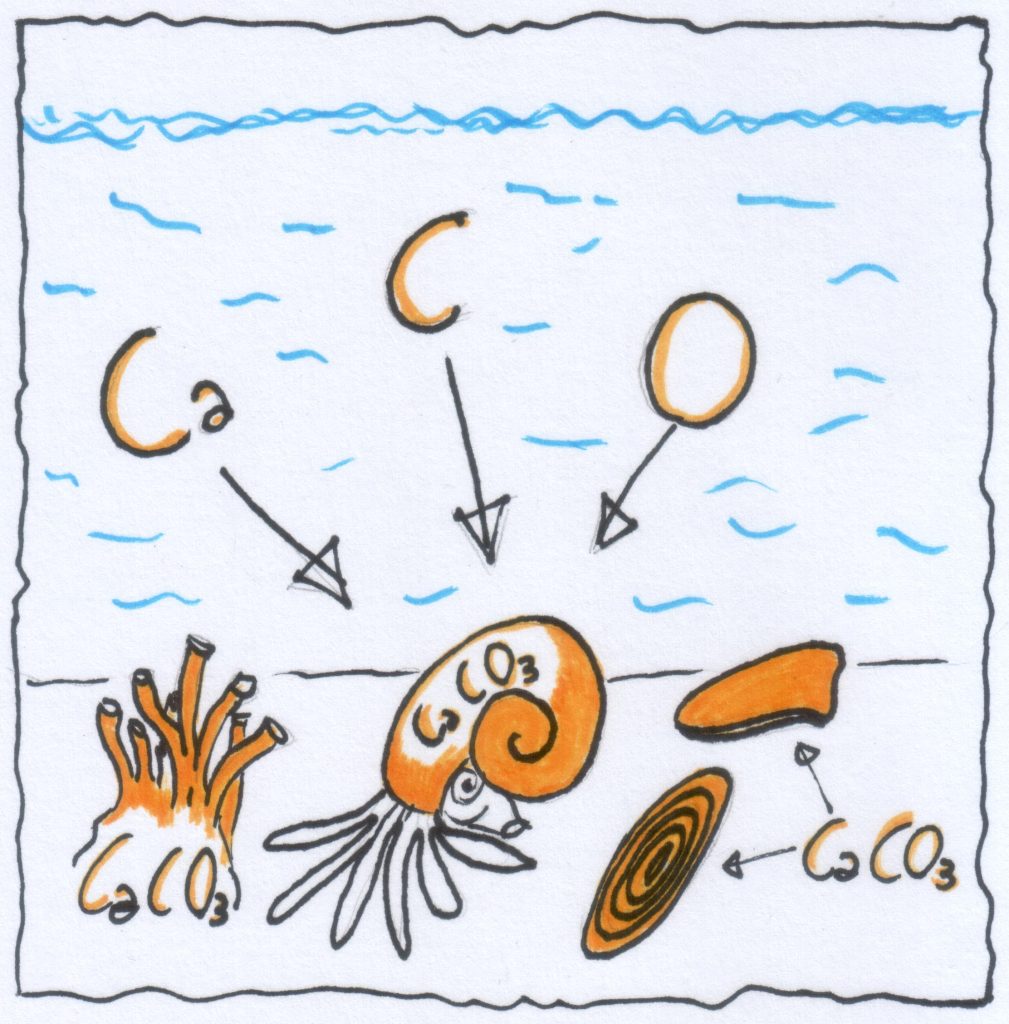
La litogenesi organogena
Gli organismi formano le rocce carbonatiche (…e silicee)
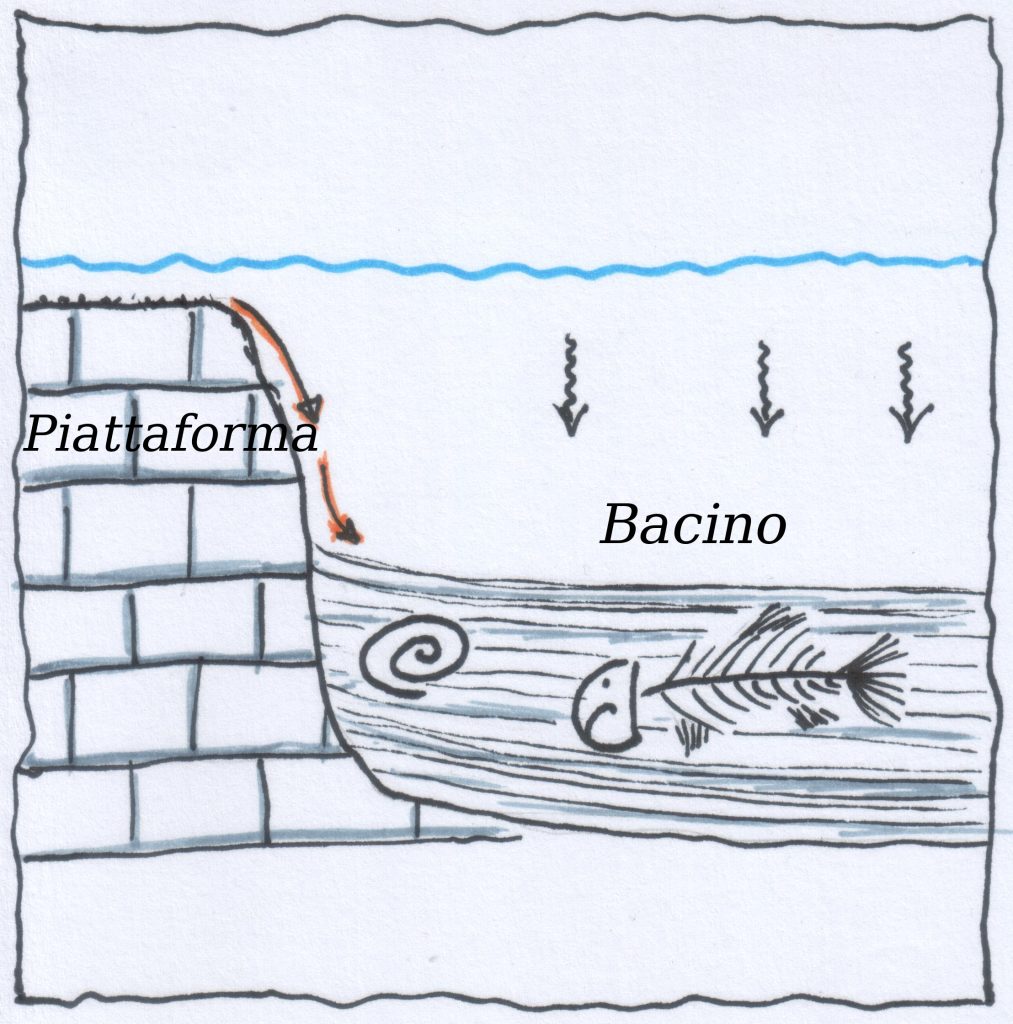
Al di sotto di una certa profondità (ccd) la calcite non riesce più depositarsi salvo eccezioni, ma viene interamente disciolta. Anche per l’aragonite esiste la acd meno profonda. La silice invece potrà depositarsi anche oltre la ccd.
La concentrazione di CO2 è direttamente responsabile della profondità della ccd. Se aumenta la [CO2], diminuisce la profondità della ccd!
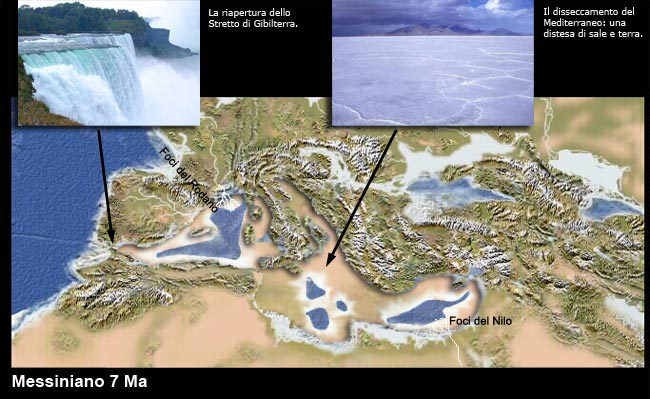
Le evaporiti
Un bacino chiuso sulle coste del Mar Caspio
Molto in generale, in un bacino chiuso o semichiuso nel quale l’evaporazione superi l’apporto di acqua si depositeranno gesso, cloruri, carbonati…
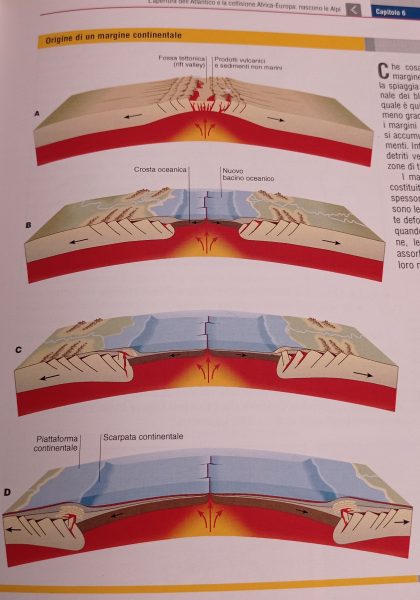
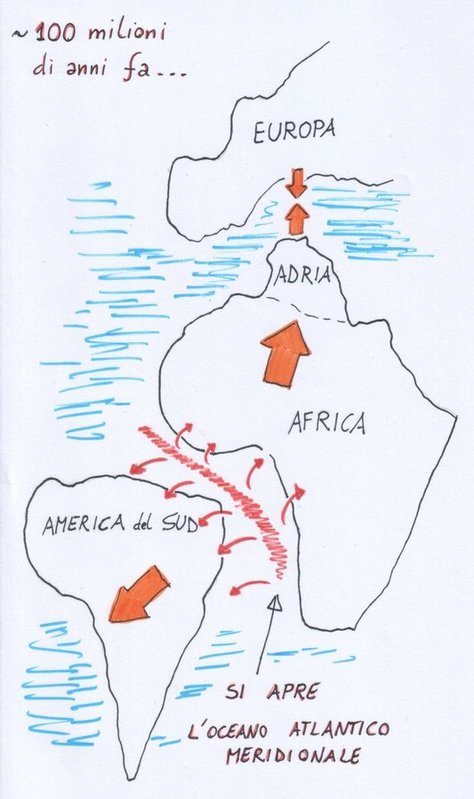
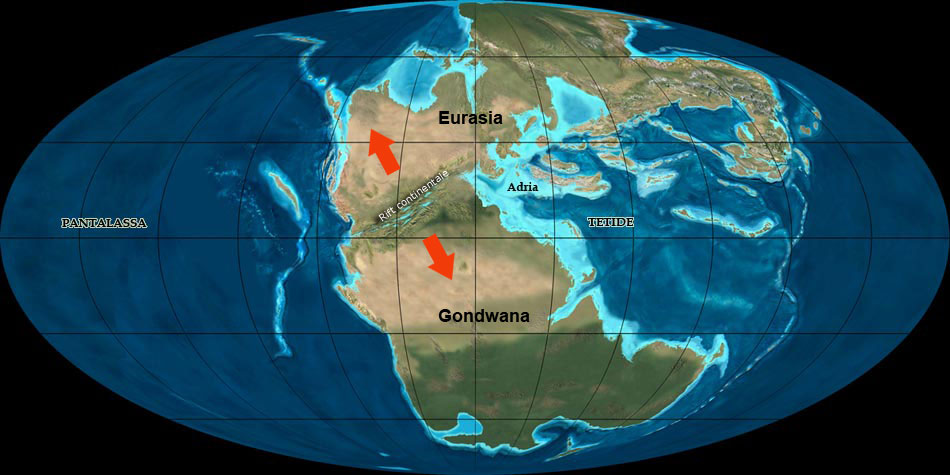
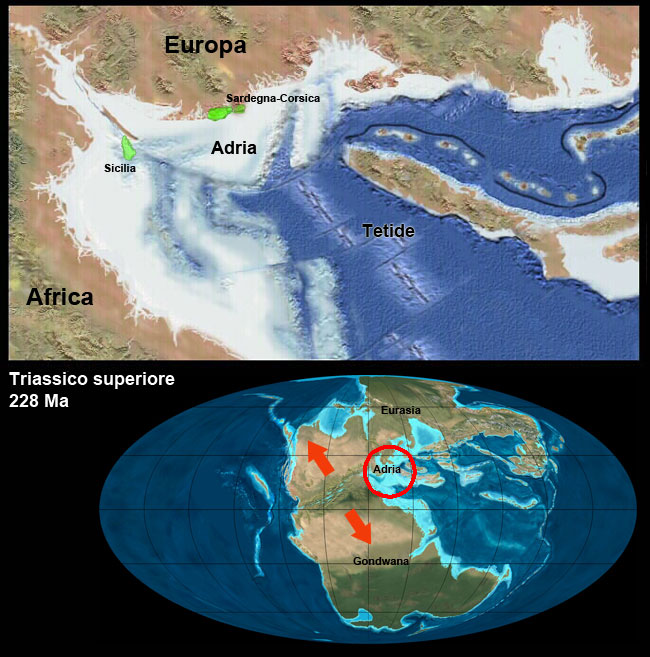
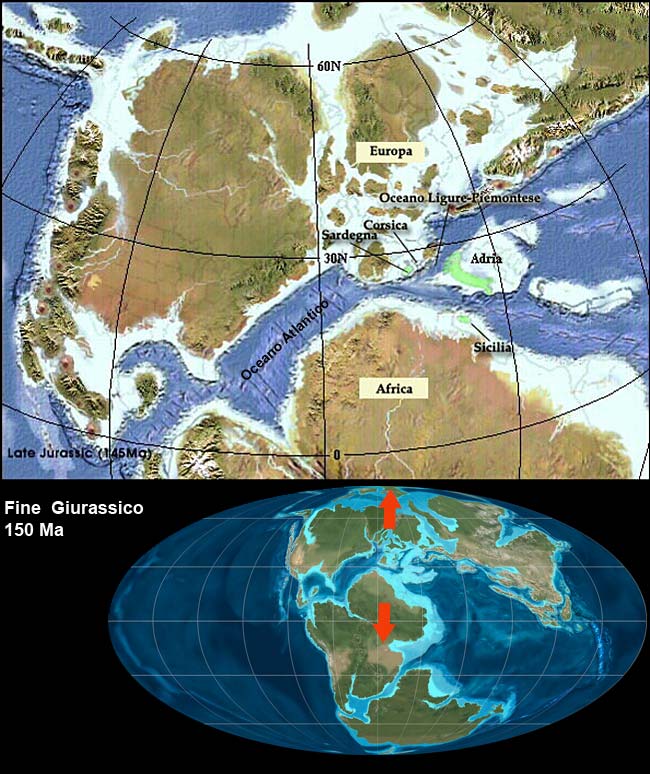
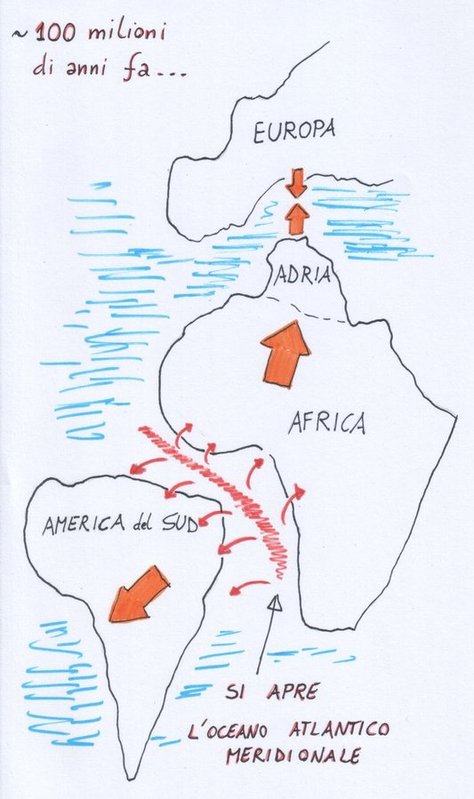
Nuova crosta terrestre viene formata e poi riassorbita per subduzione, nuove catene montuose si formano, gli oceani si aprono e si chiudono.
Mentre questi processi agiscono, la forma del rilievo viene modificata continuamente da numerosi agenti. Vediamone alcuni:
I processi gravitativi per primi (quando qualcosa viene sollevata, se manca l’equilibrio tende a sempre a cadere!).
Immagine da Wikipedia
Esistono molti tipi di frane. Qualche esempio:
Mentre nei casi precedenti sono le onde a scavare e destabilizzare un terreno che probabilmente si sta sollevando per movimenti crostali, nel caso successivo è il torrente a scavare e ad aver scavato provocando il crollo del terreno incoerente e probabilmente anche zuppo di acqua.
Mari, fiumi e acque superficiali sono agenti erosivi.
Anche i ghiacciai che sembrano fermi sono agenti di modellamento del rilievo:
Le rocce possono anche sciogliersi.
Il fenomeno del Carsismo agisce sia in superficie (karren, campi solcati, doline), sia nel sottosuolo.
Tutto cambia in continuazione, che lo vediamo o no, inesorabilmente, sempre!
Grazie!
Ste!
We use cookies to personalise content and ads, to provide social media features and to analyse our traffic. We also share information about your use of our site with our social media, advertising and analytics partners.
Cookies settings
Accept
Privacy & Cookie policy
Privacy & Cookies policy
Cookies list
Cookie name
Active
Privacy Policy
What information do we collect?
We collect information from you when you register on our site or place an order.
When ordering or registering on our site, as appropriate, you may be asked to enter your: name, e-mail address or mailing address.
What do we use your information for?
Any of the information we collect from you may be used in one of the following ways:
To personalize your experience
(your information helps us to better respond to your individual needs)
To improve our website
(we continually strive to improve our website offerings based on the information and feedback we receive from you)
To improve customer service
(your information helps us to more effectively respond to your customer service requests and support needs)
To process transactions
Your information, whether public or private, will not be sold, exchanged, transferred, or given to any other company for any reason whatsoever, without your consent, other than for the express purpose of delivering the purchased product or service requested.
To administer a contest, promotion, survey or other site feature
To send periodic emails
The email address you provide for order processing, will only be used to send you information and updates pertaining to your order.
How do we protect your information?
We implement a variety of security measures to maintain the safety of your personal information when you place an order or enter, submit, or access your personal information.
We offer the use of a secure server. All supplied sensitive/credit information is transmitted via Secure Socket Layer (SSL) technology and then encrypted into our Payment gateway providers database only to be accessible by those authorized with special access rights to such systems, and are required to?keep the information confidential.
After a transaction, your private information (credit cards, social security numbers, financials, etc.) will not be kept on file for more than 60 days.
Do we use cookies?
Yes (Cookies are small files that a site or its service provider transfers to your computers hard drive through your Web browser (if you allow) that enables the sites or service providers systems to recognize your browser and capture and remember certain information
We use cookies to help us remember and process the items in your shopping cart, understand and save your preferences for future visits, keep track of advertisements and compile aggregate data about site traffic and site interaction so that we can offer better site experiences and tools in the future. We may contract with third-party service providers to assist us in better understanding our site visitors. These service providers are not permitted to use the information collected on our behalf except to help us conduct and improve our business.
If you prefer, you can choose to have your computer warn you each time a cookie is being sent, or you can choose to turn off all cookies via your browser settings. Like most websites, if you turn your cookies off, some of our services may not function properly. However, you can still place orders by contacting customer service.
Google Analytics
We use Google Analytics on our sites for anonymous reporting of site usage and for advertising on the site. If you would like to opt-out of Google Analytics monitoring your behaviour on our sites please use this link (https://tools.google.com/dlpage/gaoptout/)
Do we disclose any information to outside parties?
We do not sell, trade, or otherwise transfer to outside parties your personally identifiable information. This does not include trusted third parties who assist us in operating our website, conducting our business, or servicing you, so long as those parties agree to keep this information confidential. We may also release your information when we believe release is appropriate to comply with the law, enforce our site policies, or protect ours or others rights, property, or safety. However, non-personally identifiable visitor information may be provided to other parties for marketing, advertising, or other uses.
Registration
The minimum information we need to register you is your name, email address and a password. We will ask you more questions for different services, including sales promotions. Unless we say otherwise, you have to answer all the registration questions.
We may also ask some other, voluntary questions during registration for certain services (for example, professional networks) so we can gain a clearer understanding of who you are. This also allows us to personalise services for you.
To assist us in our marketing, in addition to the data that you provide to us if you register, we may also obtain data from trusted third parties to help us understand what you might be interested in. This ‘profiling’ information is produced from a variety of sources, including publicly available data (such as the electoral roll) or from sources such as surveys and polls where you have given your permission for your data to be shared. You can choose not to have such data shared with the Guardian from these sources by logging into your account and changing the settings in the privacy section.
After you have registered, and with your permission, we may send you emails we think may interest you. Newsletters may be personalised based on what you have been reading on theguardian.com. At any time you can decide not to receive these emails and will be able to ‘unsubscribe’.
Logging in using social networking credentials
If you log-in to our sites using a Facebook log-in, you are granting permission to Facebook to share your user details with us. This will include your name, email address, date of birth and location which will then be used to form a Guardian identity. You can also use your picture from Facebook as part of your profile. This will also allow us and Facebook to share your, networks, user ID and any other information you choose to share according to your Facebook account settings. If you remove the Guardian app from your Facebook settings, we will no longer have access to this information.
If you log-in to our sites using a Google log-in, you grant permission to Google to share your user details with us. This will include your name, email address, date of birth, sex and location which we will then use to form a Guardian identity. You may use your picture from Google as part of your profile. This also allows us to share your networks, user ID and any other information you choose to share according to your Google account settings. If you remove the Guardian from your Google settings, we will no longer have access to this information.
If you log-in to our sites using a twitter log-in, we receive your avatar (the small picture that appears next to your tweets) and twitter username.
We are in compliance with the requirements of COPPA (Childrens Online Privacy Protection Act), we do not collect any information from anyone under 13 years of age. Our website, products and services are all directed to people who are at least 13 years old or older.
Updating your personal information
We offer a ‘My details’ page (also known as Dashboard), where you can update your personal information at any time, and change your marketing preferences. You can get to this page from most pages on the site – simply click on the ‘My details’ link at the top of the screen when you are signed in.
Online Privacy Policy Only
This online privacy policy applies only to information collected through our website and not to information collected offline.
Your Consent
By using our site, you consent to our privacy policy.
Changes to our Privacy Policy
If we decide to change our privacy policy, we will post those changes on this page.


















{kind=link}
{kind=link}